
Öga



Ett öga (latin: oculus) är ett anatomiskt organ för att förnimma ljus. Olika typer av ljuskänsliga organ finns i nästan hela djurriket. De enklaste varianterna särskiljer bara om omgivningen är mörk eller ljus. Mer komplexa ögon används för att ge ett synsinne. Många komplexa organismer, såsom däggdjur, fåglar, reptiler och fiskar har två ögon som är placerade i samma plan och vars intryck tolkas som en enda, tredimensionell "bild", liksom hos människan. Andra djurarter, till exempel kaniner och kameleonter har ögonen i olika plan och får två separata bilder.
Ögontyper
[redigera | redigera wikitext]Hos de flesta ryggradsdjur och vissa blötdjur fungerar ögat genom att projicera bilder på en ljuskänslig näthinna (retina). Signaler skickas därifrån till hjärnan via synnerven. Sådana ögon är ofta ungefär sfäriska och fyllda med en genomskinlig geleartad substans som kallas glaskropp, har en lins som fokuserar ljuset, och en iris som reglerar hur mycket ljus som kommer in i ögat.
Ögon hos bläckfiskar, fiskar, amfibiska djur och ormar har ofta en fast linsform och fokuserar blicken genom att teleskopera ögat (på samma sätt som en kamera fokuserar).
Arter som skiljer sig mycket åt kan ha väldigt olika typer av ögon, men de tenderar att likna varandra i funktion och utseende när de är fullt utvecklade. Blötdjurens ögon verkar till exempel ha utvecklats från andra organ än ryggradsdjurens ögon, och kan vara ett exempel på hur evolutionen lett till samma slutresultat. Ryggradsdjurens ögon utvecklades från hjärnceller under den embryoniska tiden, medan blötdjurens ögon växte in från hudceller. Ryggradsdjurens näthinnor har lager med neuroner framför de ljuskänsliga cellerna, medan blötdjurens näthinnor har de ljuskänsliga cellerna framför neuronerna, och har därför ingen blind fläck och möjligen skarpare syn, men också långsammare bildåterhämtning från näthinnan och därför sämre sinne för rörelser. Vissa huvudfotningar har ingen fysisk lins, utan en väldigt liten ljusöppning (ungefär som en camera obscura). Vissa blötdjur har en konkav spegel för att fokusera ljuset tillsammans med en lins.
Fasettögon finns hos leddjur, och ger en pixel-baserad bild (inte flera bilder som många tror). Varje sensor har sin egen lins och ljuskänsliga celler. Vissa ögon har upp emot 28 000 sådana sensorer, arrangerade i ett hexagonalt nät, vilket kan ge ett 360-gradigt synfält. Fasettögon är väldigt känsliga för rörelser. Vissa leddjurs fasettögon har ett fåtal fasetter var med en näthinna som kan skapa en bild, vilket ger en syn baserad på flera bilder, alla ur olika vinklar, sammansmälta till en bild med mycket hög upplösning.
Djurrikets troligen mest komplicerade ögon är fasettögonen hos stomatopoder, mantisräkor, en grupp bland kräftdjuren. Färgseendet är mycket avancerat med det största kända antalet färgreceptorer, och de kan se i 12 färgkanaler (jfr 3 hos människan), s.k. hyperspektralt seende. Varje enskilt öga har även djupseende. Vidare kan de se polariserat ljus och bedöma polarisationsplanet, och de kan också se ultraviolett och kanske infrarött ljus.
Trilobiterna, som nu är utdöda, hade unika fasettögon med genomskinliga kalk-kristaller som linser. (De flesta andra leddjur har mjuka ögon.) Antalet linser i sådana ögon varierar - vissa hade bara en, andra hade flera tusen linser per öga.
Några av de enklaste ögonen återfinns hos djur såsom sniglar, och kan inte se i den vardagliga betydelsen. De har ljuskänsliga celler, men ingen lins och inget annat sätt att projicera en bild på de cellerna. De kan särskilja mellan ljust och mörkt (dag och natt) men inte mer. Det gör att sniglar kan undvika direkt solljus.
Ögats evolutionära utveckling
[redigera | redigera wikitext]
Hur en så komplex struktur som ögats projicering skulle ha kunnat utvecklats av sig självt sägs ofta vara en svår fråga för evolutionsteorin. Darwin behandlade ämnet i sin Om arternas uppkomst genom att hävda att det inte var så konstigt om de mest primitiva varianterna också hade en funktion, och därefter muterade lite i taget.
Forskarna Dan-Eric Nilsson och Susanne Pelger i Lund har visat genom teoretiska beräkningar[1][2] att ett primitivt optiskt sinnesorgan skulle kunna ha utvecklats till ett komplext människolikt öga på en rimlig tid (mindre än en miljon år), enbart genom små mutationer och naturliga urvalsprocesser.
Ögon i olika djurarter visar att de har anpassats till sina omgivningar. Till exempel har rovfåglar skarpare blick än människor, och vissa rovfåglar som jagar på dagen kan se ultraviolett ljus. Dessutom visar de parallella utvecklingarna av ryggradsdjursögon och blötdjursögon att det inte är konstigt att ögat har utvecklats genom evolution.
Anatomi
[redigera | redigera wikitext]
Hos en nyfödd människa är ögats diameter omkring 17 mm och hos en vuxen människa är ögongloben omkring 25 mm i diameter. Ögat slutar växa ungefär i 6–7-årsåldern.
Däggdjursögon är konstruerade för att fokusera ljus på näthinnan. Alla delar som ljuset färdas genom innan det når näthinnan är glasklart genomskinliga för att förhindra en förlust i ljusstyrka innan det når näthinnan. Hornhinnan i kombination med linsen ser till att ljusstrålarna fokuseras på näthinnan. Ljuset orsakar kemiska förändringar i de ljuskänsliga cellerna i näthinnan, som aktiveras och skickar nervimpulser till hjärnan.
Ljuset, som kommer in i ögat via ett yttre medium såsom luft eller vatten, passerar först hornhinnan och vidare in i den främre ögonkammaren. Hornhinnan, som är rundad, står för den huvudsakliga (2/3) brytningen av ljuset. Den främre ögonkammaren är fylld med kammarvätska, en helt klar vätska som till sin sammansättning liknar blodserum. Trycket från kammarvätskan spänner ut hornhinnan så att den blir helt konvex, vilket är nödvändigt för att ljuset ska samlas på linsen. Den främre ögonkammaren avgränsas bakåt av iris, en ring av i huvudsak lucker bindväv och stråk av glatt muskulatur. I iris finns gott om melanocyter, som producerar färgämnet melanin. Mängden melanin avgör ögonfärgen. Mitt i iris finns ett hål, pupillen. Pupillens storlek regleras av en ringmuskel (m. sphincter pupillae) och ett radialt muskelstråk (m. dilator pupillae). Pupillen fungerar som bländaren på en kamera, och ser till att ljusnivån i ögat hålls konstant. Om för mycket ljus släpps in skulle näthinnan skadas, och om för lite ljus släpps in ser ögat inget. Innanför irisen finns den bakre ögonkammaren. Denna begränsas bakåt av linsen, en konvex, fjädrande skiva som fokuserar ljuset på näthinnan. Innan ljuset når näthinnan måste det ta sig genom glaskroppen, en geléartad struktur som fyller ut ögats insida. Glaskroppens uppgift är framförallt att stadga upp ögat, men även att bryta ljuset.
Linsen är via tunna trådar fäst vid utskott i corpus ciliare, som omsluter den ringformiga ciliarmuskeln. För att se ett föremål som befinner sig långt borta, slappnar ciliarmuskeln av och får större diameter, vilket leder till att linsen dras ut och blir plattare. När ciliarmuskeln drar ihop sig fjädrar linsen tillbaka till en tjockare, mer konvex form. När vi åldras, förlorar linsen gradvis sin förmåga att fjädra tillbaka, och det leder till att det blir svårt att fokusera på näraliggande föremål. Det finns flera brytningsfel som kommer av hornhinnan och linsens form, och från ögats längd, till exempel översynthet, närsynthet, och astigmatism.
Omkring glaskroppen finns tre lager av vävnader:
Ytterst finns tunica externa bulbi, som delas in i tre delar, senhinnan (sclera), gränsregionen (limbus) och hornhinnan (cornea). Senhinnan hjälper till att hålla ögats form och förhindrar oönskad ljusgenomträngning. Utseendemässigt är senhinnan normalt vit, tjockast kring synnerven och tunnast vid ögonmusklernas fästen och i området där synnerven passerar (lamina cribrosa). Senhinnan består av bindväv innehållande framför allt kollagen.
I mitten finns tunica vasculosa bulbi eller uvea, som även den delas in i tre delar, regnbågshinnan (iris), strålkroppen (corpus ciliare, ciliarkroppen) och åderhinnan (choroidea, koroidea). Melanocyter i åderhinnan ger ögats inre dess mörka färg, som förhindrar att det bildas störande reflexer i ögat. Åderhinnan innehåller också kapillärer, som levererar syre och näring till näthinnans fem yttersta lager och fraktar bort restprodukter. Näthinnans fem innersta lager får näring från retinas centrala artär som kommer in i ögat tillsammans med synnerven och vars kärlförgreningar ligger i näthinnans nervfiberlager. Näthinnan innehåller även pigmentepitel (med melanin) och de ljuskänsliga tapparna och stavarna samt nerver.
Innerst finns tunica interna bulbi, som delas in i näthinnans seende och inte seende del. Gränsen mellan dessa två heter ora serrata.
För att maximera ljusabsorptionen är näthinnan slät. Den har dock två punkter som är annorlunda: blinda fläcken, den punkt där synnerven går in, och i centrum gula fläcken (macula lutea) med bäst synskärpa. I denna finns centralgropen (fovea centralis), som är en liten grop klädd enbart med tappar.
Tappar och stavar
[redigera | redigera wikitext]
Näthinnan innehåller två typer av ljuskänsliga celler: tappar och stavar. Trots att de har samma uppbyggnad och metabolism, har de väldigt olika funktioner.
Stavarna är mycket ljuskänsliga, vilket gör att de fungerar även i mycket svagt ljus. Det är de här cellerna som gör att människor och djur kan se i exempelvis månljus. Dock kan de inte skilja mellan olika färger, och de har dålig synskärpa (det vill säga de har svårt att skilja på detaljer). Det är därför som saker verkar få mindre färg, ju mörkare omgivningen blir.
Tapparna å andra sidan ger hög synskärpa under goda ljusförhållanden. Ju tätare tapparna sitter, desto högre blir synskärpan. Olika sorters tappceller reagerar också på olika färger (våglängder av ljus), vilket gör dessa ansvariga för en organisms färgseende. Tapparna har även en möjlighet att bli trötta efter ett intensivt synintryck. Om man till exempel tittar intensivt på ett rött streck på marken så ser man ett grönt streck om man tittar på en vit yta. Det beror på att tapparna blir trötta och de sänder inte lika mycket röda signaler längre, då verkar den delen av synfältet mer grön (alltså motsatsfärgen) än resten av synfältet och därför ser man då ett grönt streck.
Hos däggdjur och fåglar med god syn finns det ofta ett eller flera områden i ögats näthinna med extra mycket tappar. Hos människan (och en del andra djur) finns denna i en rund, lite tunnare fördjupning av näthinnan. Denna grop kallas för gula fläcken[3] eller fovea (fullständigt latinskt namn: fovea centralis, centrala gropen) och sitter rakt bakom linsen. En del djur har en horisontellt formad fovea vilket gör att deras detaljseende fungerar bra längs med hela horisonten. Många fåglar har två foveor som även innehåller mycket fler tappar än hos exempelvis människan och som därmed ger dem ytterligare skarpare syn.
Eftersom tapparna behöver mycket ljus för att fungera optimalt blir det problem för exempelvis astronomer, då de inte kan se på ljussvaga stjärnor med ögats vanliga fokus, där ljuset inte är tillräckligt för att stimulera tapparna. Därför betraktar ofta astronomer stjärnorna genom "ögonvrån" (genom att titta lite bredvid), där andelen ljuskänsligare stavar är högre.
Både tappar och stavar är alltså känsliga för ljus, men för ljus av olika frekvenser. De innehåller båda ett pigmenterat ljusreceptor-protein, som i stavarna heter rhodopsin, i tapparna iodopsin. Både tappar och stavars ljusreceptorprotein består av en proteindel (stavar: opsin, tappar: fotodopsin), som är associerad med retinal, som inte är ett protein utan syntetiseras från vitamin A i näthinnans pigmentepitel. Processen som ljusreceptorproteinerna genomgår är likartade - när proteinet utsätts för elektromagnetisk strålning av en särskild våglängd och intensitet (det vill säga ljus inom det synliga spektret) bryts retinalet ned från sin normala konfiguration (11-cis-retinal) till en isoform (transretinal). Retinalen släpper också från opsinet/fotodopsinet. Denna process startar en signalväg som stänger jonkanaler i cellmembranet vilket förorsakar en impuls som så småningom når hjärnans syncentrum.
I närmare detalj fungerar rhodopsinet/iodopsinet i princip som en så kallad G-protein-kopplad receptor, vars aktivering leder till att ett enzym, cGMP-fosfodiesteras, börjar omvandla signalmolekylen cGMP till 5'-GMP. Då cGMP behövs för öppning av natriumkanalerna leder spjälkning till stängning vilket ger en hyperpolarisering av cellen. Denna hyperpolarisering leder till att utsöndringen av neurotransmittorer till synapsen avbryts. Detta kan verka bakvänt, men i näthinnans fotoreceptorer har neurotransmittorerna en inhiberande effekt på synapsen, och utsöndras normalt konstant. Att de slutar utsöndras leder på så sätt till aktivering av synapsen.
Flera sensoriska celler är kopplade till samma bipolära nervcell, som sedan är kopplad till en enda ganglie (nervknut) som skickar informationen vidare till syncentrat. Men tapparna i fovea är ofta kopplade individuellt till de bipolära cellerna och behöver sällan dela ganglie. Ju flera sensoriska celler som delar ganglie desto mindre skarp blir bilden från den delen av näthinnan.
Enligt den trikromatiska färgteorin uttyds färger genom att iodopsinet i tapparna finns i olika varianter. En typ bryts ner av den specifika ljusvåglängd som kommer från rött ljus, en från grönt ljus och en från blått ljus, medan den fjärde typen av tappar är känslig för ultraviolett ljus. Människan och andra högre utvecklade apor har tre typer av tappar som främst reagerar på blått, grönt och rött. De flesta andra däggdjur har två typer av tappar som främst regarera på blått och grönt medan fåglar har fyra typer av tappar så att de förutom blått, grönt och rött också kan se ultraviolett ljus. Sköldpaddor har sex olika typer av tappar. Om alla tre typer stimuleras lika mycket, ser man vitt, och om ingen stimuleras ser man svart. Oftast stimuleras de olika typerna olika mycket, vilket leder till att man ser olika färger. De tre färgerna kallas primärfärger. Om man blandar två av dem får man sekundärfärger, och blandar man två sekundärfärger får man tertiärfärger, och så vidare. Felfunktion hos någon av tapptyperna ledar till olika grad av färgblindhet. För rovfåglarna blir det mer komplicerat.
Ögats rörelser
[redigera | redigera wikitext]
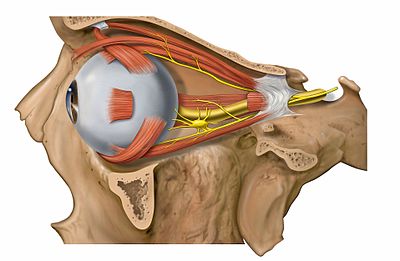 |
Det finns sex yttre ögonmuskler med ursprung från ögonhålan som fäster på ögonbulbens sidor. Dessa styr rörelserna av ögat och håller det kvar i ögonhålan. Fyra av de sex musklerna är raka muskler (rectus), som något förenklat vrider ögat i varsin riktning. Musculus rectus lateralis vrider ögat lateralt, m. rectus medialis vrider ögat medialt, m. rectus superior och inferior vrider ögat uppåt respektive nedåt. De två kvarvarande musklerna är de sneda ögonmusklerna, m. obliquus superior och m. obliquus inferior. M. obliquus superior löper genom en ligamentring, trochlea, i anteromediala ögonhåletaket. Från denna går sedan muskeln bakåt och fäster på laterala och posteriora sidan av ögat. Vid kontraktion av muskeln kommer ögat då att vridas inåt och nedåt. M. obliquus inferior utgår från främre delen av ögonhålan och går bakåt för att fästa posterolateralt. Rörelsen blir då vridning utåt och uppåt. Tre av de raka ögonmusklerna, m. rectus superior, inferior och medialis) samt den undre sneda ögonmuskeln (m. obliquus inferior) innerveras av den tredje (III) kranialnerven, nervus oculomotorius, medan m. obliquus superior innerveras av fjärde kranialnerven (IV), nervus trochlearis, och m. rectus lateralis av sjätte kranialnerven (VI), nervus abducens. Bilden till höger visar även m. levator palpebrae superioris, som lyfter det övre ögonlocket, samt ganglion ciliare, som styr pupillkontraktion och ackommodation.
Ögats skyddsmekanismer
[redigera | redigera wikitext]Ögat är omgivet av korta hårstrån som kallas ögonfransar. De är till för att skydda ögat mot vattendroppar, damm och andra partiklar som skulle kunna komma in i ögat. Varje öga har tårkörtlar. De sänder kontinuerligt ut vätska som sprids ut över ögat när man blinkar. Detta förhindrar att ögat torkar ut. Om ögonfransarna misslyckats med sin uppgift och släppt in smuts i ögat, producerar tårkörtlarna extra mycket vätska för att skölja rent och då blinkar man samtidigt för att pressa ut smutsen.
Ögonproblem och -sjukdomar
[redigera | redigera wikitext]Se separat artikel om ögonsjukdomar samt sammanställningen i Kategori:Ögonsjukdomar.
Se även
[redigera | redigera wikitext]Källor
[redigera | redigera wikitext]- Kandel, Schwartz, Jessell "Principles of Neural Science", 4/e, McGraw-Hill: Health Professions Division, 2000, ISBN 0-07-112000-9
- Malm, Liedholm "Akut neurologi" (1986), 7:e upplagan 2004, ISBN 91-631-4784-X
- ^ ”Eye design and evolution” (på engelska). Lunds universitet. Arkiverad från originalet den 26 februari 2009. https://web.archive.org/web/20090226224013/http://www.lu.se/o.o.i.s/9103. Läst 15 januari 2009.
- ^ Nilsson och Pelger (1994). ”A pessimistic estimate of the time required for an eye to evolve” (på engelska). Proceedings of the Royal Society B 256: sid. 53-58. http://www.jstor.org/pss/49593. Läst 15 januari 2009.
- ^ Remington, Lee Ann (2005) (på engelska). Clinical Anatomy of the visual system. Elsevier. ISBN 978-0-7506-7490-4
|